Вегетативная регуляция мочевого пузыря
Содержание статьи
6.5. ВЕГЕТАТИВНАЯ ИННЕРВАЦИЯ МОЧЕВОГО ПУЗЫРЯ И ПРЯМОЙ КИШКИ — Клиника лечения боли — Pain Clinic
Мочеиспускание осуществляется согласованной деятельностью m. sphincter pupillae и m. detrusor pupillae.
Происходит это при взаимодействии соматической и вегетативной нервной систем.
Мочевой пузырь имеет двойную вегетативную (симпатическую и парасимпатическую) иннервацию.
Спинальный парасимпатический центр находится в боковых рогах спинного мозга на уровне сегментов S2-S4 (ядро Онуфа). От него парасимпатические волокна идут в составе тазовых нервов и иннервируют гладкие мышцы мочевого пузыря, преимущественно детрузор. Парасимпатическая иннервация обеспечивает сокращение детрузора и расслабление сфинктера, что обеспечивает опорожнение мочевого пузыря.
Симпатическую иннервацию осуществляют волокна от боковых рогов спинного мозга (сегменты L1-L2), далее они проходят в составе подчеревных нервов (nn. hypogastrici) к внутреннему сфинктеру мочевого пузыря. Симпатическая стимуляция приводит к сокращению мышцы пузырного треугольника, что предотвращает рефлюкс мочи в мочевой пузырь при мочеиспускании.
Функционирование мочевого пузыря обеспечивается спинальным рефлексом: сокращение сфинктера сопровождается расслаблением детрузора — мочевой пузырь наполняется мочой. Когда он наполнился, сокращается детрузор и расслабляется сфинктер, моча выводится. По такому типу мочеиспускание осуществляется у детей в первые годы жизни, когда акт мочеиспускания не контролируется сознательно, а осуществляется механизмом безусловного рефлекса.
У здорового взрослого человека мочеиспускание осуществляется по типу условного рефлекса: человек сознательно может задержать мочеиспускание при возникновении позыва и опорожнять пузырь по желанию. Произвольная регуляция осуществляется с участием корковых сенсорных и моторных зон. К супраспинальным контролирующим механизмам относится мостовой центр (Барингтона), входящий в состав ретикулярной формации. Афферентная часть этого условного рефлекса начинается рецепторами, которые находятся в участке внутреннего сфинктера. Далее сигнал через спинномозговые узлы, задние корешки, задние канатики, продолговатый мозг, мост, средний мозг направляется к сенсорному участку коры (girus fornicatus), откуда по ассоциативным волокнам импульсы поступают в корковый двигательный центр мочеиспускания, который локализируется в парацентральной дольке (lobulus paracentralis).
Эфферентная часть рефлекса в составе корково-спинномозгового пути проходит в боковых и передних канатиках спинного мозга и заканчивается в спинальных центрах мочеиспускания (S2-S4 сегменты), которые имеют двустороннюю корковую связь. Далее волокна через передние корешки, половое сплетение и половой нерв (п. pudendus) достигают внешнего сфинктера мочевого пузыря. При сокращении внешнего сфинктера детрузор расслабляется и наступает торможение позыва к мочеиспусканию. При мочеиспускании напрягается не только детрузор, но и мышцы диафрагмы, брюшного пресса, в свою очередь внутренний и внешний сфинктеры расслабляются.
Нейрогенный мочевой пузырь — это синдром, который объединяет нарушения мочеиспускания, возникающие при поражении нервных путей или центров, иннервирующих мочевой пузырь и обеспечивающих функцию произвольного мочеиспускания. При двустороннем поражении коры и ее связей со спинальными (крестцовыми) центрами мочеиспускания возникают нарушения мочеиспускания по центральному типу, которые могут проявляться полной задержкой мочи (retention urinae), возникающей в острый период болезни (миелит, спинальная травма и т. п.). В этом случае угнетается рефлекторная деятельность спинного мозга, исчезают спинальные рефлексы, в частности, рефлекс опорожнения мочевого пузыря — сфинктер находится в состоянии сокращения, детрузор расслаблен и не функционирует. Моча растягивает пузырь до больших размеров. В таких случаях необходима катетеризация мочевого пузыря. В дальнейшем (через 1-3 нед) повышается рефлекторная возбуждаемость сегментарного аппарата спинного мозга и задержка мочи сменяется ее недержанием. Моча выделяется периодически небольшими порциями по мере того, как она накапливается в пузыре; т. е. мочевой пузырь опорожняется автоматически, функционирует по типу безусловного (спинального) рефлекса: накопление определенного количества мочи приводит к расслаблению сфинктера и сокращению детрузора. Такое нарушение мочеиспускания называется периодическое (перемежающееся) недержание мочи (incontinention intermittens).
Если патологический процесс локализируется в крестцовых сегментах спинного мозга, корешках конского хвоста и периферических нервах (n. hypogastricus, n. pudendus), т. е. нарушается парасимпатическая иннервация мочевого пузыря, возникают нарушения функции тазовых органов по периферическому типу. В острый период болезни в результате паралича детрузора и сохранения эластичности шейки мочевого пузыря возникает полная задержка мочи, или парадоксальная задержка мочи (ishuria paradoxa) с выделением мочи каплями при переполненном мочевом пузыре в случае задержки мочеиспускания (за счет механического перерастяжения сфинктера мочевого пузыря). В последующем шейка мочевого пузыря теряет свою эластичность, а сфинктер в таком случае раскрытый, наступает денервация внутреннего и внешнего сфинктеров, поэтому возникает истинное недержание мочи (incontinention vera) с выделением мочи по мере поступления ее в мочевой пузырь.
Вегетативная иннервация прямой кишки и её сфинктеров осуществляется по типу иннервации мочевого пузыря. Различие состоит в том, что в прямой кишке нет мышцы детрузора, а его роль выполняют мышцы брюшного пресса.
Источник
Нейрональный контроль мочеиспускания. Часть 1.
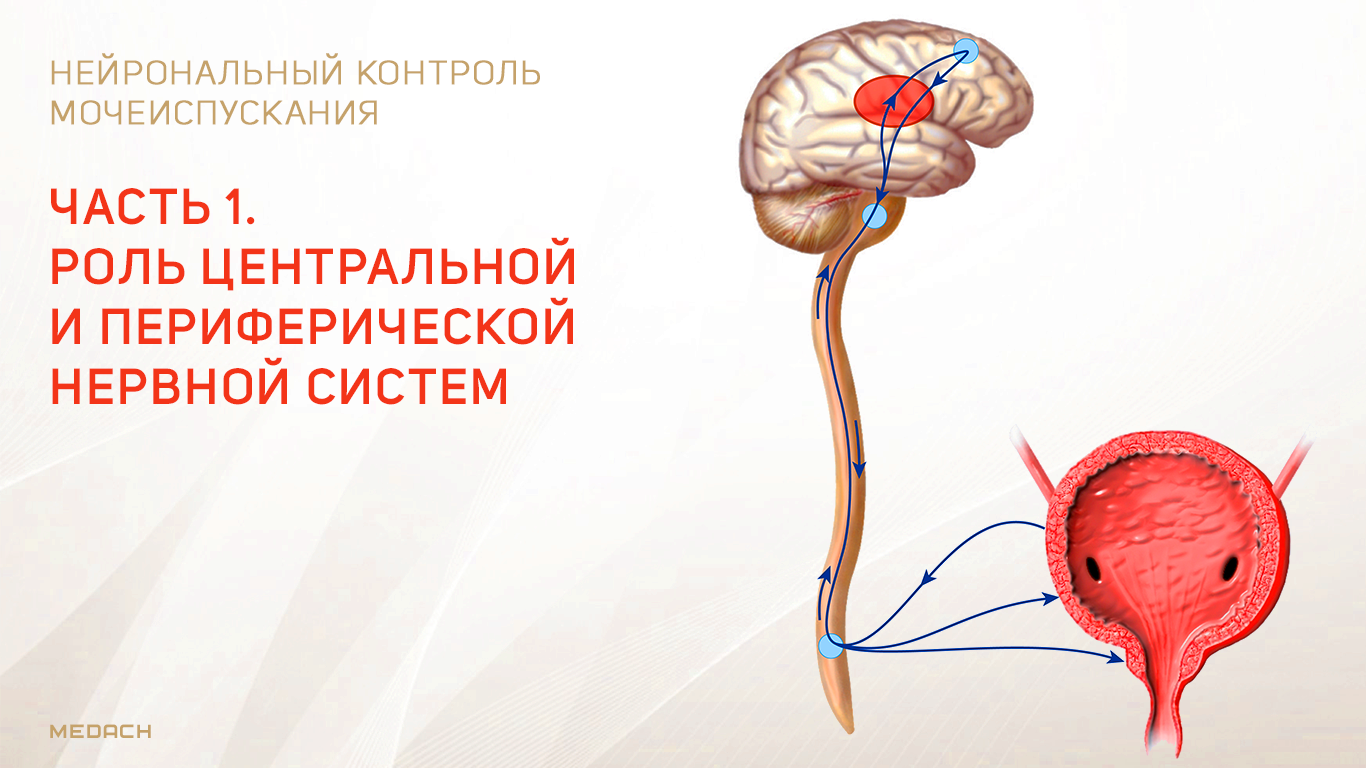
Хранение мочи и периодическое опорожнение мочевого пузыря (МП) зависят от координированной работы гладких и поперечнополосатых мышц, условно разделенных на два структурно функциональных отдела: резервуара (мочевой пузырь) и выпускной системы, состоящей из шейки МП, уретры и наружного сфинктера мочеиспускательного канала. За координированную работу вышеперечисленных структур отвечают сложные нейронные системы управления, расположенные в периферических ганглиях, спинном и головном мозге.
Обеспечение сознательного контроля мочеиспускания требует сложных взаимодействий между вегетативным (симпатика и парасимпатика) и соматическим отделами нервной системы.
Симпатические нервные волокна берут свое начало из боковых рогов грудопоясничного отдела (спинномозговой центр Якобсона) и ганглиях нижнего брыжеечного сплетения и достигают МП в составе подчревного нерва. Симпатические постганглионарные нервы выделяют норадреналин, который активирует β-адренергические рецепторы, ингибирующие сокращение детрузорной мышцы, и α-адренергические рецепторы, возбуждающие мускулатуру уретры и шейки МП. Симпатическое воздействие приводит к расслаблению мышцы, изгоняющей мочу, сокращению мускулатуры уретры и шейки МП и ингибированию интрамуральных ганглиев МП.
Центральный отдел парасимпатической иннервации МП располагается в промежуточных ядрах крестцовых сегментов. Холинергические преганглионарные волокна из промежуточных ядер посылают свои аксоны через тазовые нервы к ганглиозным клеткам тазового сплетения и интрамуральным нейронам в стенке мочевого пузыря. Ганглиозные клетки в свою очередь возбуждают детрузор мочевого пузыря, что приводит к его сокращению с последующим опорожнением. Основными медиаторами парасимпатической нервной системы тут являются ацетилхолин и другие нехолинергические медиаторы. Ацетилхолин действует опосредованно, возбуждая M3-холинорецепторы клеток детрузора. Парасимпатические нервные окончания в нервно-мышечных синапсах и в парасимпатических ганглиях тоже имеют холинорецепторы, возбуждение этих рецепторов на нервных окончаниях может усиливать (через рецепторы М1) или подавлять (через рецепторы М4) высвобождение медиаторов в зависимости от интенсивности нейронного возбуждения. Основной нехолинергический медиатор это АТФ, который активирует внутриклеточную пуринергическую систему через возбуждение P2X рецепторов и тоже способствует сокращению детрузора. Парасимпатические волокна вызывают расслабление ГМК уретры путем высвобождения оксида азота (NO).
Аксоны соматических двигательных нейронов передних рогов крестцовых сегментов S2-S4 (ядро Онуфа) проходят в срамном нерве и иннервируют поперечно-полосатые мышцы наружного уретрального сфинктера. Нейроны более медиально расположенного моторного ядра на том же уровне позвоночника иннервируют мускулатуру тазового дна.
Афферентные пути НМП состоят из цепочек чувствительных нейронов. Первые нейроны, располагающиеся в спинальных ганглиях на уровне S2-S4 и T11-L2, реагируют на пассивное растяжение и активное сокращение мышц МП и передают эту информацию на нейроны второго и третьего порядка. Эти нейроны обеспечивают координированную работу спинальных рефлексов и восходят к вышележащим структурам головного мозга, контролирующих фазы накопления и опорожнения МП. Наиболее важные афферентные волокна от мочевого пузыря идут в составе тазового нерва, в то время как чувствительность от шейки МП и уретры передается по срамным и подчревным нервам. Афферентные волокна этих нервов состоят из миелинизированных (Аδ) и немиелинизированных (С) аксонов. Aδ-волокна передают информацию о наполнении мочевого пузыря. С-волокна нечувствительны к изменению объема мочевого пузыря в физиологических условиях, поэтому они называются «тихими». Они реагируют главным образом на патологические стимулы, такие как химическое раздражение или охлаждение.
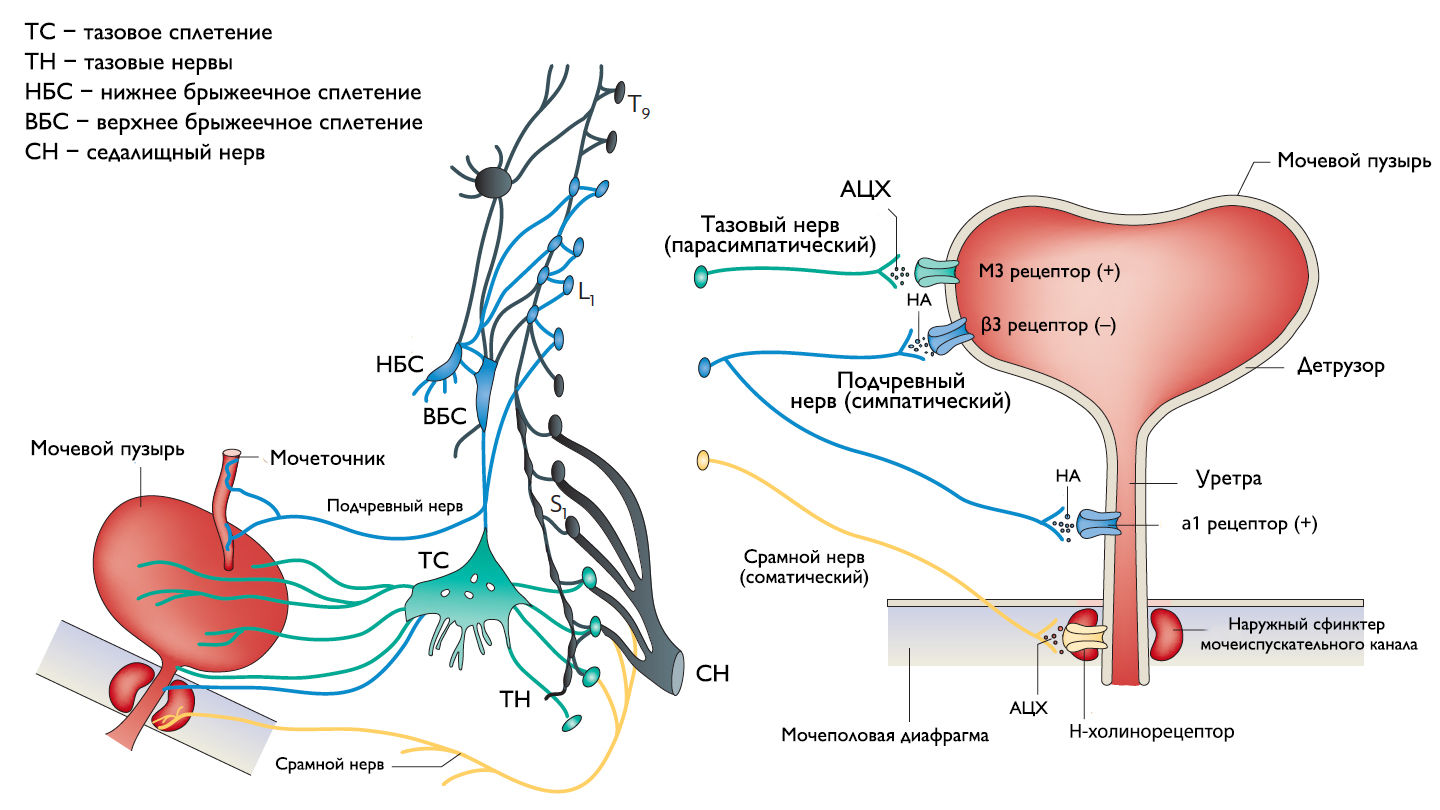
Рисунок 1 | Иннервация нижних мочевыводящих путей.
Специфический и неспецифический восходящий супраспинальный сенсорный путь
Одни спинномозговые промежуточные нейроны посылают восходящие волокна к определенным областям моста и среднего мозга, участвующим в мочеиспускании. Другие промежуточные нейроны передают информацию из нижних мочевыводящих путей в структуры переднего мозга, включая таламус и гипоталамус. Спиноталамический и спиногипоталамический тракты хотя и не играют главную роль в мочеиспускании, но могут включаться в сознательный контроль полноты мочевого пузыря. Чувствительные зоны коры ГМ через спиноталамический тракт информируются о состоянии наполнения МП.
Мостовой центр мочеиспускания (МЦМ) и его нисходящие спинномозговые двигательные пути
Впервые центр управления мочеиспусканием был открыт в дорсальной части моста Баррингтоном в 1925 году и с тех пор называется мостовым центром мочеиспускания или ядром Баррингтона. МЦМ располагается в области покрышки моста. Нейроны МЦМ имеют нисходящие возбуждающие синаптические контакты с клетками парасимпатических преганглионарных мотонейронов, иннервирующих постганглионарные клетки мочевого пузыря. Электрическая и химическая стимуляция МЦМ у крыс и кошек инициирует сокращение мочевого пузыря и расслабляет сфинктер уретры имитируя нормальное мочеиспускание. Цикл рефлекса мочеиспускания состоит из трех фаз, контролируемых различными центральными механизмами: фаза реализации безопасной среды — для начала мочеиспускания человеку необходимо осознание, что окружающие обстановка комфортна; фаза релаксации наружного уретрального сфинктера; и фаза сокращения мышцы, выталкивающей мочу. Процесс нормального мочеиспускания невозможен без какой-либо из этих фаз. МЦМ является командным центром мочеиспускания, который контролирует последовательное переключение фазы расслабления наружного уретрального сфинктера на фазу сокращения детрузорной мышцы.
Мостовой центр удержания мочи (МЦУ) и его нисходящие спинномозговые двигательные пути
Его роль заключается в расслаблении детрузора и сокращении наружного уретрального сфинктера. МЦУ располагается вентролатеральней МЦМ. Синапсы волокон МЦУ возбуждают ядро Онуфа в крестцовых сегментах спинного мозга, повышая таким образом тонус наружного сфинктера уретры. Стимуляция области МЦУ останавливает мочеиспускание, возбуждает мышцы тазового дна и сокращает уретральный сфинктер. Наоборот, двусторонние поражения МЦУ вызывают недержание мочи, чрезмерную детрузорную активность, невозможность хранения мочи и снижение тонуса уретрального сфинктера. На сегодняшний день нет анатомических доказательств связи между МЦУ и МЦМ, и было высказано предположение, что эти центры функционально независимы.
Роль кортикальных областей
Наиболее частые симптомы поражения кортикальных областей ГМ это поллакиурия и ургентное недержание мочи. Поэтому Andrew и Nathan выдвинули гипотезу, что отсоединение лобной или передней поясной извилины от гипоталамуса приводит к непроизвольному началу мочеиспускания [3]. Действительно, префронтальная кора головного мозга человека и передняя поясная извилина активируются во время мочеиспускания [4].
Мозжечок и базальные ганглии
Существует ряд исследований о том, что мозжечок и базальные ганглии оказывают в основном ингибирующее действие на мочевой пузырь. Мозжечковая патология приводит к увеличению частоты мочеиспускания и ургентному недержанию мочи. Симптомы гиперактивного мочевого пузыря также встречаются при болезни Паркинсона. Поскольку нет прямых связей этих областей с МЦМ, ингибирующее влияние, вероятно, косвенное через структуры переднего и среднего мозга.
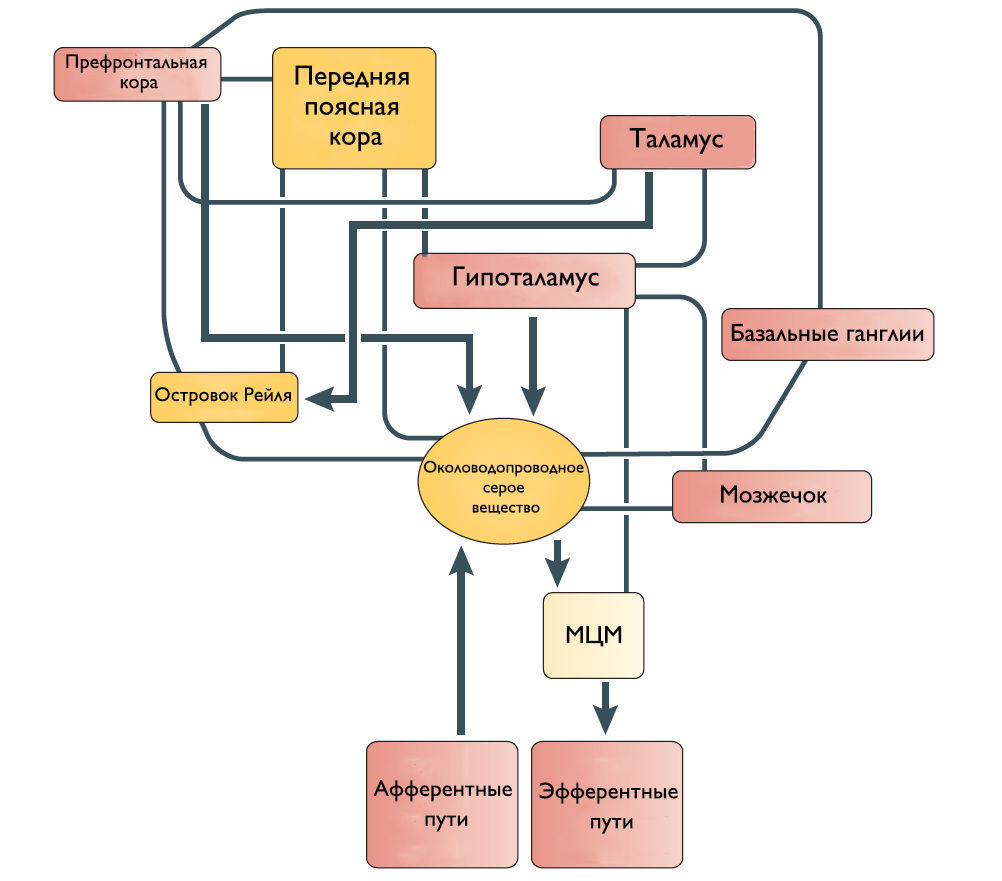
Рисунок 2 | Предположительное схематическое изображение связей между различными структурами переднего мозга и ствола мозга, которые участвуют в контроле мочеиспускания.
- Liao L., Madersbacher H. (ed.). Neurourology: Theory and Practice. — Springer, 2019.
- Clare J. Fowler et al. The neural control of micturition. — Nature Reviews | Neuroscience, volume 9. — June 2008.
- Andrew J, Nathan PW. Lesions of the anterior frontal lobes and disturbances of micturition and defecation. Brain. 1964;87:233-62.
- Griffiths, Derek J. «Use of al imaging to monitor central control of voiding in humans.» Urinary Tract. Springer, Berlin, Heidelberg, 2011. 81-97.
Нашли опечатку? Выделите фрагмент и нажмите Ctrl+Enter.
Источник
2.5.2. Иннервация мочевого пузыря. Расстройства мочеиспускания.
Мышечная основа мочевого пузыря и прямой кишки состоит преимущественно из гладкой мускулатуры, а поэтому иннервируется вегетативными волокнами. В то же время в состав пузырного и анального сфинктеров входят поперечнополосатые мышцы, что дает возможность произвольного их сокращения и расслабления.
В акте мочеиспускания можно выделить 2 компонента:
непроизвольно-рефлекторный;
произвольный.
Рефлекторное опорожнение мочевого пузыря осуществляется благодаря сегментарным центрам симпатической (боковые рога спинного мозга на уровне сегментов L1-L3) и парасимпатической иннервации (сегменты S2 — S4). Симпатическая иннервация осуществляется нижним подчревным сплетением, пузырными нервами, а парасимпатическая иннервация — тазовым нервом. Симпатические волокна сокращают сфинктер и расслабляют детрузор (гладкие мышцы). Парасимпатические волокна вызывают расслабление сфинктера и сокращение детрузора. При повышении тонуса симпатической нервной системы возникает задержка мочи, а возбуждение парасимпатического центра приводит к опорожнению мочевого пузыря.
Поперечнополосатая мускулатура тазовых органов (наружный сфинктер мочевого пузыря) иннервируется половым нервом (S2 — S4). Чувствительные волокна от наружного сфинктера уретры направляются в сегменты S2 — S4, где и замыкается рефлекторная дуга. Другая часть волокон через систему боковых и задних канатиков направляется к коре больших полушарий. Связи спинальных центров с корой (парацентральная долька и верхние отделы передней центральной извилины) прямые и перекрестные. Кора больших полушарий обеспечивает произвольный акт мочеиспускания. Корковые центры не только регулируют произвольное мочеиспускание, но и могут затормаживать этот акт.
Регуляция мочеиспускания представляет собой своеобразный циклинческий процесс. Наполнение мочевого пузыря приводит к раздражению рецепторов, находящихся в детрузоре, слизистой оболочке пузыря и проксимальной части уретры. От рецепторов импульсы передаются как в спинной мозг, так и в вышестоящие отделы — диэнцефальную область и кору больших полушарий головного мозга. Благодаря этому формируется чувство позыва на мочеиспускание. Пузырь опорожняется в результате координированного действия нескольких центров: возбуждение спинномозгового парасимпатического, некоторого угнетения симпатического, произвольного расслабления наружного сфинктера и активного напряжения мышц брюшного пресса. После завершения акта мочеиспускания начинает преобладать тонус симпатического спинномозгового центра, способствующего сокращению сфинктера, расслаблению детрузора и наполнению пузыря. При соответствующем наполнении цикл повторяется.
Таблица 4
Основные нарушения мочеиспускания
Тип нарушения мочеиспускания | Клинические проявленияя | Очаг поражения в нервной системе |
центральный | императивные позывы задержка мочи недержание мочи | Поражение проводящих корково-спинномозговых путей |
периферический | парадоксальная ишурия истинное недержание мочи | Поражение парасимпатического спинномозгового центра Поражение парасимпатического и симпатического спинномозговых центров |
Задержка мочи возникает при спазме сфинктера, слабости детрузора. Но она обычно имеет преходящий характер. Временная задержка мочи сменяется недержанием мочи вследствие «растормаживания» спинальных сегментарных центров. Это недержание является, по существу, автоматическим, непроизвольным опорожнением мочевого пузыря по мере его наполнения и называется периодическим недержанием мочи. При этом вследствие сохранения рецепторов и чувствительных путей ощущение позыва к мочеиспусканию приобретает характер императивного (больной должен помочиться немедленно, иначе произойдет непроизвольное опорожнение мочевого пузыря).
Парадоксальная ишурия возникает при переполнении мочевого пузыря, когда сфинктер частично раскрывается под давлением мочи, и моча выделяется каплями.
Истинное недержание мочи, паралич мочевого пузыря характеризуется тем, что моча постоянно выделяется каплями, по мере ее поступления в мочевой пузырь
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Источник
Вегетативная нервная система человека
Вегетативная (автономная) нервная система — отдел нервной системы, регулирующий деятельность внутренних органов, желез внутренней и внешней секреции, кровеносных и лимфатических сосудов.
Вегетативная нервная система иннервирует весь организм, все органы и ткани. Деятельность вегетативной нервной системы не зависит от воли человека. Однако все вегетативные функции подчиняются центральной нервной системе, в первую очередь — коре больших полушарий.
Функции:
нервная регуляция функций всех органов и тканей организма (кроме скелетных мышц);
регуляция обмена веществ;
поддержание гомеостаза организма;
приспособительные реакции всех позвоночных.
Особенности вегетативной нервной системы:
очаговое расположение в мозге вегетативных нервных центров;
эффекторные (двигательные) нейроны расположены за пределами центральной нервной системы в узлах вегетативных нервных сплетений;
двухнейронный эфферентный нервный путь от мозга до рабочего органа;
преобладают немиелинизированные нервные волокна, т.е. скорость проведения нервных импульсов ниже, чем в соматической нервной системе.
строение вегетативной нервной системы
Анатомически и функционально вегетативная нервная система подразделяется на симпатическую, парасимпатическую и метасимпатическую.
Все структуры и системы организма иннервируются волокнами вегетативной нервной системы. Отделы вегетативной нервной системы находятся в относительном функциональном антагонизме, обеспечивая автоматическую регуляцию органов и систем без участия сознания человека.
Важнейшие органы имеют двойную иннервацию. Полые внутренние органы имеют тройную (симпатическую, парасимпатическую и метасимпатическую) иннервацию.
В симпатическом и парасимпатическом отделах имеются центральная и периферическая части.
Центральную часть вегетативной нервной системы образуют вегетативные ядра — тела нейронов, лежащих в спинном и головном мозге. Они осуществляют координацию работы всех трех частей вегетативной нервной системы.
Периферическую часть вегетативной нервной системы образуют отходящие от ядер нервные волокна, вегетативные ганглии, лежащие за пределами центральной нервной системы, и нервные сплетения в стенках внутренних органов.
Симпатические и парасимпатические центры находятся под контролем коры больших полушарий и гипоталамуса.
| строение и особенности | симпатический отдел | парасимпатический отдел |
| центральный отдел | Ядра в боковых рогах спинного мозга:
| 4 ядра в стволе головного мозга:
Ядра во II — IV сегменте крестцового отделе спинного мозга |
| периферический отдел | парный симпатический ствол; нервные сплетения; нервы | нервные узлы в стенках внутренних органов или рядом с органами; нервы |
| медиаторы | норадреналин | ацетилхолин |

Симпатический отдел вегетативной нервной системы
Симпатические ядра расположены в спинном мозге на уровне грудных позвонков. Отходящие от ядер нервные волокна заканчиваются за пределами спинного мозга в симпатических узлах, расположенных по бокам позвоночника. От них берут начало нервные волокна, которые подходят ко всем органам.
Симпатическая нервная система усиливает обмен веществ, повышает возбуждаемость большинства тканей, мобилизует силы организма на активную деятельность.
Симпатический отдел возбуждается при воздействии адреналина.
параСИМПАТИЧЕСКИЙ ОТДЕЛ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Парасимпатические ядра лежат в продолговатом мозге и в крестцовой части спинного мозга. Нервные волокна от ядер продолговатого мозга входят в состав блуждающих нервов. От ядер крестцовой части нервные волокна идут к кишечнику, органам выделения. Парасимпатические нервные узлы располагаются в стенках внутренних органов или возле органов.
Парасимпатическая система способствует восстановлению израсходованных запасов энергии, регулирует работу организма во время сна.
Парасимпатический отдел нервной системы возбуждается под воздействием ацетилхолина.
метаСИМПАТИЧЕСКИЙ ОТДЕЛ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Метасимпатическая нервная система представлена нервными сплетениями и мелкими ганглиями в стенках пищеварительного тракта, мочевого пузыря, сердца и некоторых других органов.
Функция: осуществляет связь между внутренними органами (минуя головной мозг); местные вегетативные рефлексы..
Известно, что многие внутренние органы, извлеченные из организма, продолжают выполнять присущие им функции. Например, сохраняется перистальтическая и всасывательная функция тонкой кишки. Такая относительная функциональная независимость объясняется наличием в стенках этих органов метасимпатического отдела вегетативной нервной системы.
Особенности метасимпатического отдела нервной системы:
Обладает собственным нейрогенным ритмом и имеет полный набор необходимых для самостоятельной рефлекторной деятельности звеньев: чувствительный, вставочный и эффекторный нейрон с соответствующим медиаторным обеспечением.
Имеет собственные сенсорные элементы (механо-, хемо-, термо-, осморецепторы), которые посылают в свои внутренние сети информацию о состоянии иннервируемого органа, а также способны передавать сигналы в ЦНС.
Ограничена: охватывает только некоторые внутренние органы.
Не имеет своего центрального аппарата; ее связь с ЦНС осуществляется нейронами симпатического и парасимпатического отделов.
Существование специальных местных метасимпатических механизмов регуляции функций имеет определенный физиологический смысл. Их наличие увеличивает надежность регуляции функций. Эта регуляция может происходить в случае выключения связи с центральными структурами. При этом ЦНС освобождается от избыточной информации.
Органы с разрушенными метасимпатическими путями утрачивают способность к координированной моторной деятельности и другим функциям.
Влияние симпатического и парасимпатического отделов на отдельные органы
Симпатический отдел:
повышает частоту и силу сердечных сокращений;
стимулирует выброс адреналина;
повышает уровень глюкозы в крови;
повышает артериальное давление;
вызывает расширение артерий головного мозга, легких и коронарных артерий;
угнетает перистальтику кишечника и работу пищеварительных желез (в том числе слюнных), сокращает гладкомышечные сфинктеры;
угнетает перистальтику мочеточников, расслабляет мускулатуру и сокращает сфинктер мочевого пузыря;
расширяет бронхи и бронхиолы, усиливает вентиляцию легких;
расширяет зрачки.
Парасимпатический отдел:
уменьшает частоту и силу сердечных сокращений;
понижает уровень глюкозы в крови;
снижает артериальное давление;
усиливает перистальтику кишечника и стимулирует работу пищеварительных желез (в том числе слюнных), расслабляет гладкомышечные сфинктеры;
усиливает перистальтику мочеточников, сокращает мускулатуру и расслабляет сфинктер мочевого пузыря;
сужает бронхи и бронхиолы, уменьшает вентиляцию легких;
сужает зрачки.
РЕГУЛЯЦИЯ РАБОТЫ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Все механизмы регуляции деятельности внутренних органов условно объединены многоэтажной иерархической структурой.
Первый структурный уровень: внутриорганные рефлексы, имеющие метасимпатическую природу;
Второй структурный уровень: ганглии брыжеечных и солнечного (чревного) сплетений;
Оба этих низших этажа обладают отчетливо выраженной автономностью и могут осуществлять регуляцию независимо от центральной нервной системы.
Третий структурный уровень: центры спинного мозга и ствола головного мозга.
Четвертый структурный уровень: кора больших полушарий, гипоталамус, ретикулярная формация, лимбическая система и мозжечок.
Кора больших полушарий мозга: контролирует работу всех внутренних органов. Известно, что в определенных условиях у человека гипнотическим внушением можно вызвать изменение сердечного ритма, усиление потоотделения и мочеотделения, изменение метаболизма.
Рефлекторные процессы в ядерных образованиях спинного, продолговатого, среднего мозга и моста находятся под постоянным влиянием гипоталамуса.
Гипоталамические центры: поддержание гомеостаза; регуляция метаболизма; регуляция функций эндокринных желез; интеграция нервной и гуморальной регуляции вегетативных функций (через гипофиз).
Лимбическая система («висцеральный мозг»): объединение работы опорно-двигательной системы и внутренних органов: пищевое, сексуальное, оборонительное поведение, сон и бодрствование, внимание, эмоции, процессы памяти.
Мозжечок: стабилизирующее влияние на деятельность внутренних органов.
Ретикулярная формация: повышение активности нервных центров, связанных с функциями внутренних органов. Регулирует секрецию гипофизарных гормонов.
Источник