Амитотическое деление в клетках мочевого пузыря
Содержание статьи
ПРЕПАРАТ № 5 Амитоз в клетках мочевого пузыря мыши.
Мы поможем в написании ваших работ! Мы поможем в написании ваших работ! Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ? | Препарат представляет собой гистологический срез мочевого пузыря мыши, окрашенный по Нонизеду и Уиндлю (Микрофото 5). (рис. 22, 23) При малом увеличении видны: ткань построенная из клеток с хорошо различимыми оболочками и ядрами различной величины и формы (от округлой до гантелеобразной) ядра эпителиальных клеток слизистой оболочки мочевого пузыря. Слабо оксифильная цитоплазма этих клеток почти неразличима, одинаково с опущенным конденсором и прикрытой диафрагмой можно видеть клетки различной величины. Передвигая препарат, отыскиваем делящиеся клетки, где ядро сильно вытянуто в длину, в средней его части образуется перетяжка, которая истончается. Видны места, где клетка становится двуядерной, места, где есть разделение двуядерной клетки на две одноядерные (процесс цитотомии или плазмотомии). Встречаются клетки многоядерные в результате задержания цитотомии или при отсутствии ее. В крупных, неправильной округлой формы клетках содержится одно, два, три и более ядер. Среди неделящихся ядер можно видеть также ядра, находящиеся на различных стадиях амитотического деления. Эти ядра надо изучить при большом увеличении. Рис. 23. Амитоз эпителиальных клеток. Отпечаток поверхности слизистой оболочки мочевого пузыря мыши. 1 — ядро, 2 — перетяжка ядра, 3- двуядерная клетка, 4 — цитомия, 5 — многоядерные клетки. Во время прямого деления ядро остается в интерфазном состоянии и клетка не перестает функционировать. О начале амитоза свидетельствует изменение формы ядра. Основа вытягивается в длину и разрывается. Клетка становится двуядерной. В дальнейшем может произойти цитотомия. Нередко цитотомия задерживается или вообще не наступает, в результате чего возникают многоядерные клетки. Иногда видно деление ядрышка, которое удлиняется и перешнуровывается. Прямое деление клеток эпителия мочевого пузыря является дегенеративной формой, так как предшествует отмиранию этих клеток. Обозначения: 1 — делящиеся клетки, 2 — двуядерные клетки, 3 — перетяжка ядра и цитоплазмы, 4 -многоядерные клетки. Задания 1. Изучить деления клеток. 2. Изучить амитоз. Контрольные вопросы:
ЗАНЯТИЕ 3 Тема 3. ПЛАЗМАТИЧЕСКАЯ МЕМБРАНА (ПЛАЗМАЛЕММА) Содержание. Модели биологических мембран. Жидкостно-мозаичная модель. Особенности химического состава и его взаимосвязь со свойствами и функциями мембран. Клеточные стенки (оболочки). Плазмодесмы и десмосомы. Синцитий, симпласт, плазмодий. Средства наглядности. Таблицы с изображением жидкостно-мозаичной модели мембраны Оборудование и материалы. Препараты: кариотип традесканции. Таблицы с изображением кариотипов. Семена культурных злаков, луковицы, черенки ив Сосуды емкостью 250 и 1000 мл, чашки Петри, фильтровальная бумага, проволока, вода дистиллированная. Задания для аудиторной работы 1. Законспектируйте теоретическую часть занятия. 2. Ответьте на контрольные вопросы. 3. Заполните таблицу «Химический состав клеточных оболочек». Задания для внеаудиторной работы Продолжите изучение основных классов органических соединений клетки. Обратите внимание на структуру и функции углеводов, липидов и белков. Теоретическая часть Плазматическая мембрана (цитоплазматическая мембрана), или плазмалемма. Существование плазмалеммы предсказал Франц Мейен (1830), который считал, что клетка — это пространство, отграниченное от внешней среды вполне замкнутой мембраной. Овертон (начало ХХ века) установил, что плазматическая мембрана эритроцитов содержит большое количество липидов. Гортер и Грендел (1925) доказали, что мембрана состоит из двойного слоя липидов (липидный бислой). Доусон и Даниелли (1935) предложили модель бутерброда: мембрана состоит из липидного бислоя, заключенного между двумя слоями белка. Робертсон (1959) на основе анализа данных электронной микроскопии установил, что все мембраны построены по единому плану: липидный бислой + белковые молекулы. Сингер и Николсон (1972) разработали жидкостно-мозаичную модель мембраны, которая является в настоящее время общепринятой. Согласно жидкостно-мозаичной, или жидкокристаллической модели, основу мембран составляет фосфолипидный бислой. Гидрофильные глицерофосфатные части молекул фосфолипидов находятся на внешних поверхностях бислоя. Гидрофобные углеводородные части молекул фосфолипидов направлены вовнутрь бислоя. Структура бислоя поддерживается за счет поверхностного натяжения; связи между молекулами фосфолипидов называются гидрофобными. Отдельные блоки бислоя способны перемещаться относительно друг друга во всех направлениях. Кроме фосфолипидов в состав мембран входят гликолипиды и стероиды (например, холестерин). Конкретный липидный состав мембран зависит от таксономической принадлежности организмов, от тканевой принадлежности клеток и от их физиологического состояния, а также от условий обитания организмов. Белки мембран представлены простыми белками, гликопротеинами, липопротеинами, металлопротеинами и другими сложными белками. Выделяют три основных типа белков: периферические (гидрофильные, расположены на поверхности мембран), интегральные (гидрофобные, расположены в толще бислоя) и политопические (со смешанными свойствами, пронизывают мембрану насквозь). Белковые молекулы образуют непостоянные соединения между собой и небелковыми группами. В ходе химических взаимодействий конформация белков и их свойства существенно изменяются. Углеводы в составе мембран обычно представлены гликопротеинами и гликолипидами. Основная часть углеводов плазмалеммы расположена на ее внешней стороне и образует гликокаликс. Такая структура мембраны обеспечивает ее основное свойство: избирательную проницаемость. Функции плазмалеммы: барьерная, транспортная, энерготрансформирующая, информационно-сигнальная. С внешней стороны мембраны часто формируются надмембранные структуры — клеточные оболочки, или клеточные стенки. Основные функции клеточных оболочек (стенок): механическая и защитная. Основные типы клеточных оболочек: 1. У большинства низших эукариот (у водорослей, у низших грибов и грибоподобных организмов) клеточная стенка состоит из целлюлозы и гемицеллюлоз. 2. У высших грибов (Eumycota) клеточная стенка содержит хитин (реже — хитозан) и полимеры глюкозы — глюканы. У дрожжеподобных грибов хитина почти нет. 3. У высших растений первичная клеточная стенка состоит из целлюлозы. Вторичные стенки содержат суберин или лигнин. Смежные клетки разделены срединными пластинками из пектинов. У многих низших и высших растений в состав стенок входят минеральные вещества: кремнезем, известь и др. 4. У прокариот клеточная оболочка многослойная. Внутренний слой (собственно клеточная стенка) построен на основе муреина. Внешние слои имеют разнообразный химический состав. У многих видов имеется слизистая капсула из полисахаридов. |
Источник
Практическая часть
Задания
1. Изучить деления клеток.
2. Изучить мейоз.
Контрольные вопросы:
Дайте характеристику фаз редукционного деления мейоза.
Характеристика профазы, метафазы, анофазы, телофазы.
Характеристика фаз эквационного деления мейоза.
АМИТОЗ
Содержание. Деление клеток. Амитоз.
Средства наглядности. Таблицы: «Амитоз».
Оборудование и материалы.Микроскопы. Гистологические препараты.
Задания для аудиторной работы
1. Законспектируйте теоретическую часть занятия. Обратите внимание на термины, выделенные курсивом.
2. Ответьте на контрольные вопросы.
Задания для внеаудиторной работы
Продолжите изучение деления клеток.
Теоретическая часть
Амитоз- прямое деление клеток без образования хромосом этот процесс протекает без образования нитей веретена и без спирализации хромосом. Перешнуровка или расщепление сначала ядра, а затем цитоплазмы на две части с образованием двух дочерних клеток. Если цитоплазма не делится, то возникают двух — и даже многоядерные клетки. Этот способ наблюдается у эмбрионов, лейкоцитов, злокачественных опухолей. Клетки, делившиеся амитозом, не могут войти в митоз.
 Рис. 22. Амитоз эпителиальных клеток мочевого пузыря (по Нонизеду и Уиндлю)
Рис. 22. Амитоз эпителиальных клеток мочевого пузыря (по Нонизеду и Уиндлю)
Практическая часть препарат № 5 Амитоз в клетках мочевого пузыря мыши.
Препарат представляет собой гистологический срез мочевого пузыря мыши, окрашенный по Нонизеду и Уиндлю (Микрофото 5). (рис. 22, 23)
При малом увеличении видны: ткань построенная из клеток с хорошо различимыми оболочками и ядрами различной величины и формы (от округлой до гантелеобразной) ядра эпителиальных клеток слизистой оболочки мочевого пузыря. Слабо оксифильная цитоплазма этих клеток почти неразличима, одинаково с опущенным конденсором и прикрытой диафрагмой можно видеть клетки различной величины. Передвигая препарат, отыскиваем делящиеся клетки, где ядро сильно вытянуто в длину, в средней его части образуется перетяжка, которая истончается. Видны места, где клетка становится двуядерной, места, где есть разделение двуядерной клетки на две одноядерные (процесс цитотомии или плазмотомии). Встречаются клетки многоядерные в результате задержания цитотомии или при отсутствии ее. В крупных, неправильной округлой формы клетках содержится одно, два, три и более ядер. Среди неделящихся ядер можно видеть также ядра, находящиеся на различных стадиях амитотического деления. Эти ядра надо изучить при большом увеличении.
Р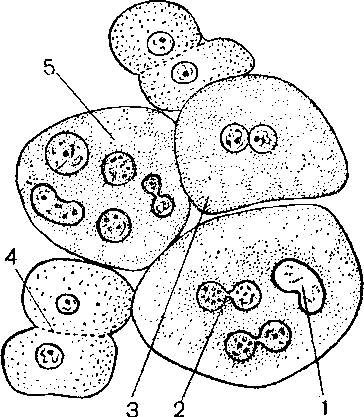 ис. 23. Амитоз эпителиальных клеток. Отпечаток поверхности слизистой оболочки мочевого пузыря мыши. 1 — ядро, 2 — перетяжка ядра, 3- двуядерная клетка, 4 — цитомия, 5 — многоядерные клетки.
ис. 23. Амитоз эпителиальных клеток. Отпечаток поверхности слизистой оболочки мочевого пузыря мыши. 1 — ядро, 2 — перетяжка ядра, 3- двуядерная клетка, 4 — цитомия, 5 — многоядерные клетки.
Во время прямого деления ядро остается в интерфазном состоянии и клетка не перестает функционировать. О начале амитоза свидетельствует изменение формы ядра. Основа вытягивается в длину и разрывается. Клетка становится двуядерной. В дальнейшем может произойти цитотомия. Нередко цитотомия задерживается или вообще не наступает, в результате чего возникают многоядерные клетки.
Иногда видно деление ядрышка, которое удлиняется и перешнуровывается. Прямое деление клеток эпителия мочевого пузыря является дегенеративной формой, так как предшествует отмиранию этих клеток.
Обозначения: 1 — делящиеся клетки, 2 — двуядерные клетки, 3 — перетяжка ядра и цитоплазмы, 4 -многоядерные клетки.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Источник
Амитоз эпителиальных клеток
При малом увеличении видны различной величины и формы ядра эпителиальных клеток слизистой оболочки мочевого пузыря. Слабо оксифильная цитоплазма этих клеток почти неразличима, однако с опущенным конденсором и прикрытой диафрагмой можно видеть клетки различной величины. В крупных, неправильной формы округлых клетках содержится одно, два, три и более ядер (Рис. 35). Среди неделящихся ядер можно видеть также ядра, находящиеся на разных стадиях амитотического деления. Эти ядра надо изучить при большом увеличении. Во время прямого деления ядро остается в интерфазном состоянии и клетка не перестает функционировать. О начале амитоза свидетельствуют изменения формы ядра. Оно вытягивается в длину, в средней части образуется перетяжка, которая быстро истончается и разрывается. Клетка становится двуядерной. В дальнейшем может произойти цитотомия. Нередко цитотомия задерживается или вообще не наступает, в результате чего возникают многоядерные клетки. Иногда видно деление ядрышка, которое удлиняется и перешнуровывается.
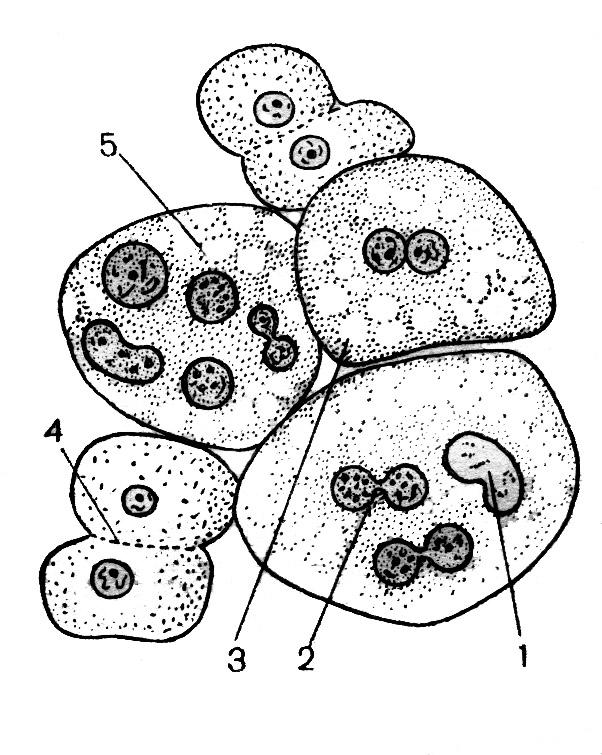
Рис. 35. Амитоз в клетках эпителия: 1 — ядро; 2 — перетяжка; 3 — двуядерная клетка; 4 — цитотомия; 5 — многоядерные клетки.
Прямое деление клеток эпителия мочевого пузыря является дегенеративной формой, так как предшествует отмиранию этих клеток.
Задание. Рассмотреть амитоз в клетках эпителия мочевого пузыря. Зарисовать и сделать все необходимые подписи к рисунку.
Пути клеточной гибели
Гибель отдельных клеток или целых групп постоянно встречается у многоклеточных организмов.
Причины гибели, процессы морфологического и биохимического характера развития клеточной смерти могут быть различны. Но они четко делятся на две категории: некроз и апоптоз (Рис. 36). Некроз связан с нарушением внутреннего гомеостаза в результате нарушения проницаемости клеточных мембран, с необратимыми изменениями митохондрий. Апоптоз — гибель клеток без их химического и физического повреждения на всех стадиях онтогенеза. Это процесс, приводящий не к лизису, не к растворению клеток, а к их фрагментации, распаду. Биологическая роль — удаление отработавших или ненужных на данном этапе клеток, а также удаление измененных или патологических клеток, особенно мутантных или зараженных вирусами.
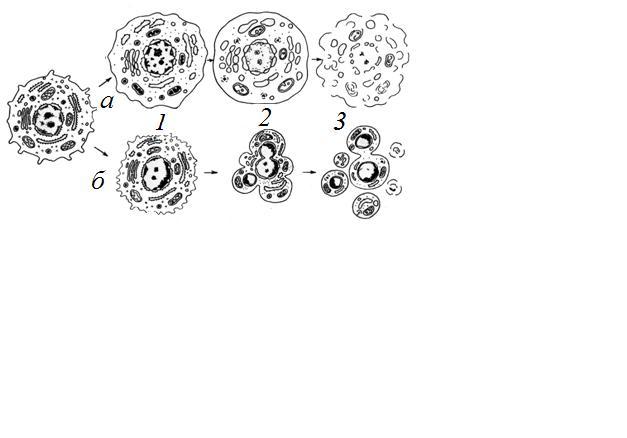
Рис. 36. Два пути клеточной гибели: а — апоптоз: 1 — специфическое сжатие клетки и конденсация хроматина; 2 — фрагментация ядра; 3 — фрагментация тела клетки на ряд апоптических телец; б — некроз: 1 — набухание клетки, вакуолярных компонентов, конденсация хроматина (кариорексис); 2 — набухание мембранных органоидов, лизис хроматина ядра (кариолизис); 3 — разрыв мембранных компонентов клетки — лизис.
Задание. Рассмотреть амитоз в клетках эпителия слизистой оболочки мочевого пузыря. Зарисовать и описать все стадии.
Эпителиальные ткани
Эпителиальные ткани — однослойные или многослойные пласты, покрывающие внутренние или внешние поверхности любого органа (Рис. 37). Развиваются: из эктодермы — эпителий кожи, нервной системы, эпителиальная выстилка переднего и заднего отделов пищеварительного тракта; из энтодермы — эпителий остальных отделов пищеварительного тракта, печени, поджелудочной железы.
Внутреннюю выстилку кровеносных сосудов образует эндотелий, который нельзя считать истинным эпителием, так как он развивается из мезодермы.
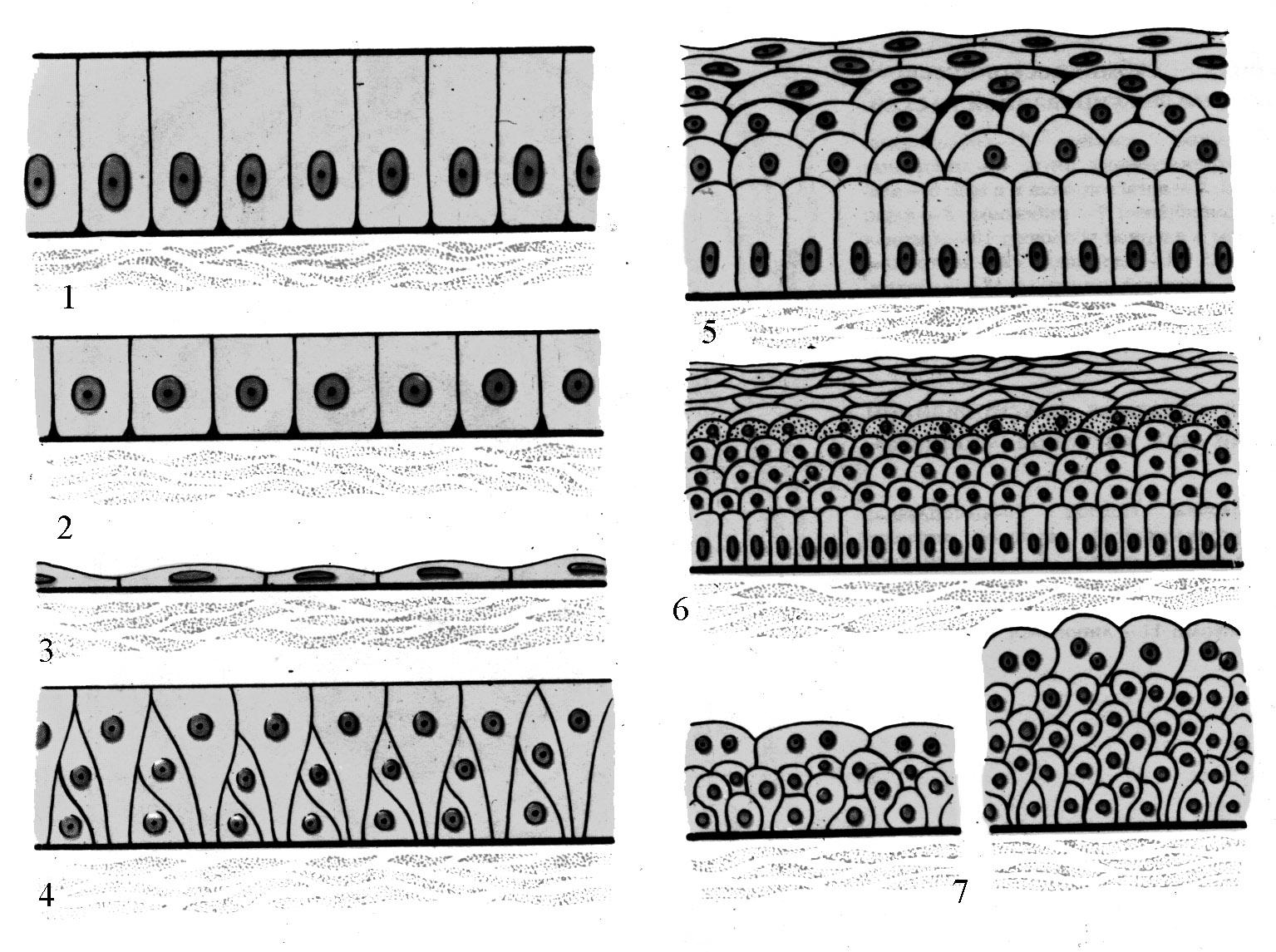
Рис. 37. Схема строения различных типов эпителия. 1- однослойный цилиндрический эпителий; 2 — однослойный кубический; 3 — однослойный плоский; 4 — многорядный эпителий; 5 — многослойный плоский неороговевающий эпителий; 6 — многослойный плоский ороговевающий; 7 — переходный эпителий в разных состояниях стенки органа
Эпителиальные клетки удерживаются между собой небольшим количеством цементирующего вещества, которое содержит гиалуроновую кислоту. Нижний слой клеток лежит на базальной мембране. Она состоит из переплетения коллагеновых волокон, которые секретируются нижележащими тканями. Так как эпителий не снабжается кровью, кислород и питательные вещества поступают в его клетки путем диффузии из лимфатических сосудов, которые разветвляются в примыкающих межклеточных пространствах.
Основная функция — защита от механических воздействий, защита от инфекций.
Эпителиальная ткань делится в зависимости от числа клеточных слоев и форм отдельных клеток на покровную и железистую. Покровный эпителий бывает простой и сложный. Простой: плоский, кубический, цилиндрический, реснитчатый и многорядный. Сложный — многослойный и переходный.
Железистый эпителий содержит секреторные клетки бокаловидной формы. Различают 2 типа секреторных клеток: экзокринные и эндокринные.
Продукты их железистых клеток могут выводиться тремя путями: 1) мерокриновый путь — секрет выделятся через клеточную мембрану на свободную поверхность без потерь цитоплазмы (потовые железы, ацинусы поджелудочной железы); 2) апокриновый путь — верхняя часть цитоплазмы отторгается при образовании секрета (в млечных железах); 3) голокриновый путь — разрушается вся клетка и секрет выталкивается через эпителий (сальные железы).
Соседние файлы в предмете Цитология
- #
- #
- #
- #
- #
- #
Источник
Новая модель дифференциации тканей мочевыводящих путей, регенерации тканей и болезней: перепрограммирование клеток простаты и клеток мочевого пузыря человека в индуцированных плюрипотентных стволовых клетках
A novel model of urinary tract differentiation, tissue regeneration, and disease: reprogramming human proe and bladder cells into induced pluripotent stem cells
Moad M, Pal D, Hepburn AC, Williamson SC, Wilson L, Lako M, Armstrong L, Hayward SW, Franco OE, Cates JM, Fordham SE, Przyborski S, Carr-Wilkinson J, Robson CN, Heer R.
Источник
Northern Institute for Cancer Re, Newcastle University, UK
Первичные модели культур животных и клеточных линий предстательной железы и развития мочевого пузыря имеют ограничения в описании биологии человека, необходимы новые стратегии, отражающие полный спектр дифференциации от плода до старения ткани. Последние достижения в области биологии показывают, что возможно прямое перепрограммирование плюрипотентных эмбриональных стволовых клеток (ЭСК) в соматические клетки. Эти клетки, называемые индуцированными плюрипотентными стволовыми клетками (ИПСК), теоретически могут регенерировать ткани простаты и мочевого пузыря взрослых, обеспечивая альтернативную стратегию изучения дифференциации.
Цель
Создание ИПСК человека, полученных из нормальной, стареющей предстательной железы человека (Про-ИПСК) и ткани мочевыводящих путей (МТ-ИПСК) и оценка их потенциала для направленной возрастной дифференцировки.
Дизайн, установки и участники
Строму простаты и мочевыводящих путей трансдуцировали генами гомеобокса 1 класса ПМ 5 (POU5F1; ранее OCT4), SRY (область определения пола Y)-контейнера 2 (SOX2), Kruppel-подобного фактора 4 (кишка) (KLF4) и вирусного гомолога онкогенеза (птичьего) v-myc миелоцитоматоза (MYC , ранее C -Myc) для генерации индуцированных плюрипотентных стволовых клеток.
Результат измерения и статистический анализ
Потенциал для дифференцировки линий простаты и мочевого пузыря сравнивали с классическими индуцированными плюрипотентными стволовыми клетками, полученными из кожи. Был использован т-тест Стьюдента.
Результаты и ограничения
Успешное перепрограммирование ткани простаты в Про-ИПСК, а ткани мочевого пузыря и мочеточников в МТ-ИПСК было продемонстрировано характерной морфологией эмбриональных стволовых клеток, экспрессией маркеров и функциональной плюрипотентностью в генерации всех трех зародышевых слоев клона.
В отличие от обычных, полученных из кожи плюрипотентных стволовых клеток, Про-ИПСК показали повышенную способность генерировать эпителиальную специфическую дифференциацию, что характеризуется рецепторами андрогенов и индукцией простат-специфического антигена. Точно так же было доказано, что МТ-ИПСК более эффективно, чем полученные из кожи ИПСК, дифференцируются в ткань мочевого пузыря, что доказывает экспрессия уротелиально-специфических маркеров: уроплакинов, клаудинов, цитокератинов и стромальных гладкомышечных маркеров: α-актина гладкой мускулатуры, калпонина и десмина.
Эти различия, скорее всего, являются эпигенетическими между отдельными линиями индуцированных плюрипотентных стволовых клеток и подчеркивают важность органоспецифических ИПСК для тканеспецифических исследований.
Выводы
Индуцированные плюрипотентные стволовые клетки представляют новую модель, которая характеризует механизмы, регулирующие дифференциацию простаты и мочевого пузыря и позволяющую разработать новые подходы к моделированию заболеваний. Регенерация клеток мочевого пузыря также представляет исключительную возможность для поступательной тканевой инженерии.
Источник